Биологи осваивают сингулярности
Повышение скорости фиксации атомно-молекулярных трансформаций позволит быстрее создавать новые лекарства. Сегодня речь идет о чтении одиночных цепей ДНК, для чего их пропускают сквозь поры.
Создан биоробот из клеток взрослого человека
Андроботы способны регенерировать даже нервные волокна. Преимущество новой технологии в том, что андроботы делаются из собственных клеток человека, это снимает опасность их отторжения иммунной системой.
Загадка происхождения черепах
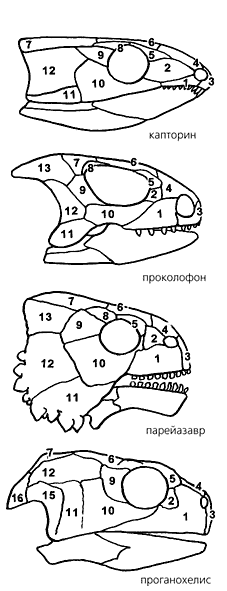
Типы фенестрации черепа рептилий: парапсидный (вверху), синапсидный (в середине) и диапсидный. Видно, что в черепе ящерицы имеется одно верхнее височное окно, у пеликозавра - нижнее, а у эвзухии - два, как и должно быть у диапсид.
Основоположником другого подхода к проблеме родства черепах стал Э.Коп, несомненно самый крупный теоретик в области палеонтологии и систематики рептилий второй половины XIX в. В 1880 г. для диадекта (Diadectus), открытого незадолго до этого в отложениях нижней перми Северной Америки, Коп установил самостоятельный отряд Cotylosauria и связал его со зверообразными ящерами. Однако дискуссии с коллегами и новые находки заставили его изменить свое мнение и обратить внимание на возможное родство котилозавров с черепахами. Позднее палеогерпетологи принялись рассматривать этот отряд столь широко, что он наполнился почти всеми рептилиеобразными животными неясного систематического положения, но со сплошной височной областью крыши черепа. В состав котилозавров вошли пермско-карбоновые сеймуриаморфы и капториноморфы, пермские парейазавры, а также пермско-триасовые проколофоны. Таким образом, черепахи, среди которых форм с височными окнами нет, попали в один ряд с древнейшими и архаичными ящерами.
Крупный шаг на пути определения родства различных пресмыкающихся сделал американский палеонтолог и систематик С.Виллистон [2]. Выделив несколько новых типов фенестрации, он установил в соответствии с ними и новые подклассы рептилий - парапсид и анапсид. В число первых Виллистон включил, например, формы с одним верхним височным окном, как у ихтиозавров и ящериц, а вторых - группы с безоконной конструкцией черепа, т.е. котилозавров (в широком смысле) и черепах. Виллистон весьма скептически относился к представлениям о родстве черепах и завроптеригий, полагая, что сходства между ними вторичны. К такой же точке зрения пришел А.Ромер и изложил ее в статьях и во всемирно известном популярном учебнике “Палеонтология позвоночных” [3].
|
|
Черепа анапсид. Такие безоконные конструкции свойственны и триасовой черепахе проганохелису, и остальным более древним пресмыкающимся, объединяемым в сборные группы котилозавров и парарептилий. Однако, как видно из нумерации, сплошная крыша черепа построена из разных наборов костей. |
Проблемы филогенетических связей черепах коснулся еще один американский палеонтолог У.Грегори [4]. Он исключил из числа ближайших черепашьих родственников завроптеригий, капториноморф, диадекта и проколофонов, оставив только парейазавров. Более того, Грегори искренне удивлялся тому, что до него этих пермских ящеров сближали лишь со зверообразными рептилиями и млекопитающими.
Сложная и запутанная классификация древнейших тетрапод заинтересовала палеонтолога Э.Олсона. Он, кстати, отмечал, что она подается его американскими коллегами, в том числе Ромером и Грегори, слишком схематично [5]. В 1947 г. Олсон предложил новый вариант системы рептилий, включив в новый подкласс - Parareptilia - все группы котилозавров, кроме капториноморф. Хотя Олсон и отмечал, что черепахи по строению черепа удивительно похожи на диадекта, в общей классификационной схеме он выводил их от основания ствола, ведущего к диадекту и парейазаврам. А в 70-х годах произошло нечто странное: Олсон отошел от некоторых своих первоначальных выводов в отношении черепах, а Ромер согласился с его исходной позицией.
- Эволюция человека
- Полиплоидия
- Питание и пища
- Первозвери
- Общая геронтология
- Ноосфера и биосфера
- Материалы
Волокна с «самосознанием»

В организме клетки «сидят» на волокнистой подложке-матриксе, препятствующей клеточной инвазии (инвазивному росту). То есть образованию метастаз. Вместе с тем в ходе внутриутробного развития клетки передвигаются. У них появляются отростки, длина которых может достигать метра (как в случае седалищного нерва).